Митохондрии. История открытия рибосом Митохондрии – энергетические центры клетки
Читайте также
Рибосомы: строение и функции
Определение 1
Замечание 1
Основной функцией рибосом является синтез белка.
Субъединицы рибосом образуются в ядрышке и потом сквозь ядерные поры отдельно друг от друга поступают в цитоплазму.
Их количество в цитоплазме зависит от синтетической активности клетки и может составлять от сотни до тысяч на одну клетку. Наибольшее количество рибосом может быть в клетках, которые синтезируют протеины. Есть они также в митохондриальном матриксе и хлоропластах.
Рибосомы различных организмов – от бактерий до млекопитающих – характеризуются подобной структурой и составом, хотя клетки прокариот имеют рибосомы меньшего размера и в большем количестве.
Каждая субъединица состоит из нескольких разновидностей молекул рРНК и десятков разновидностей белков приблизительно в одинаковой пропорции.
Маленькая и большая субъединицы находятся в цитоплазме одиночно до тех пор, пока не будут задействованы в процессе биосинтеза белка. Они объединяются друг с другом и молекулой иРНК в случае необходимости синтеза и снова распадаются, когда процесс окончен.
Молекулы иРНК, которые были синтезированы в ядре, попадают в цитоплазму к рибосомам. Из цитозоля молекулы тРНК поставляют аминокислоты к рибосомам, где с участием ферментов и АТФ синтезируются белки.
Если с молекулой иРНК соединяются несколько рибосом, то образуются полисомы , которые содержат от 5 до 70 рибосом.
Пластиды: хлоропласты
Пластиды – характерные только для растительных клеток органоиды, отсутствующие в клетках животных, грибов, бактерий и цианобактерий.
Клетки высших растений содержат 10-200 пластид. Их размер от 3 до 10 мкм. Большинство из них имеют форму двояковыпуклой линзы, но иногда могут быть в форме пластинок, палочек, зёрен и чешуек.
В зависимости от присутствующего в пластиде пигмента пигмента эти органоиды делят на группы:
- хлоропласты (гр. сhloros – зелёный) – зелёного цвета,
- хромопласты – жёлтого, оранжевого и красноватого цвета,
- лейкопласты – бесцветные пластиды.
Замечание 2
По мере развития растения пластиды одного типа способны преобразоваться в пластиды другого типа. Такое явление широко распространено в природе: изменение окраски листьев, меняется окраска плодов в процессе созревания.
Большинство водорослей вместо пластид имеют хроматофоры (обычно в клетке он один, имеет значительные размеры, имеет форму спиральной ленты, чаши, сетки или звёздчатой пластинки).
Пластиды имеют достаточно сложное внутреннее строение.
Хлоропласты имеют свои ДНК, РНК, рибосомы, включения: зёрна крахмала, капли жира. Снаружи хлоропласты ограничены двойной мембраной, внутреннее пространство заполнено стромой – полужидким веществом), которое содержит граны - особенные, свойственные лишь хлоропластам структуры.
Граны представлены пакетами плоских круглых мешочков (тилакоидов ), которые сложены как столбик монет перпендикулярно широкой поверхности хлоропласта. Тилакоиды соседних гран между собой соединяются в единую взаимосвязанную систему мембранными каналами (межмембранными ламелами).
В толще и на поверхности гран в определённом порядке расположен хлорофилл .
Хлоропласты имеют разное количество гран.
Пример 1
В хлоропластах клеток шпината содержится по 40-60 гран.
Хлоропласты не прикреплены в определённых местах цитоплазмы, а могут изменять своё положение или пассивно, или активно перемещаются ориентировано к свету (фототаксис ).
Особенно чётко активное движение хлоропластов наблюдается при значительном повышении одностороннего освещения. В таком случае хлоропласты скопляются у боковых стенок клетки, а к ориентируются ребром. При слабом освещении хлоропласты ориентируются к свету более широкой стороной и располагаются вдоль стенки клетки, обращённой к свету. При средней силе освещения хлоропласты занимают срединное положение. Таким образом достигаются наиболее благоприятные условия для процесса фотосинтеза.
Благодаря сложной внутренней пространственной организации структурных элементов хлоропласты способны эффективно поглощать и использовать лучистую энергию, а также происходит разграничение во времени и пространстве многочисленных и разнообразных реакций, составляющих процесс фотосинтеза. Реакции этого процесса, зависимые от света, происходят лишь в тилакоидах, а биохимические (темновые) реакции – в строме хлоропласта.
Замечание 3
Молекула хлорофилла очень подобна молекуле гемоглобина и отличается в основном тем, что в центре молекулы гемоглобина расположен атом железа, а не атом магния, как у хлорофилла.
В природе существует четыре типа хлорофилла: a, b, c, d.
Хлорофиллы a и b содержатся в хлоропластах высших растений и зелёных водорослей, диатомовые водоросли содержат хлорофиллы a и c, красные – a и d . Хлорофиллы a и b изучены лучше других (впервые их выделил в начале ХХ столетия российский учёный М.С. Цвет).
Кроме них существует четыре вида бактериохлорофиллов – зелёных пигментов зелёных и пурпурных бактерий: a, b, c, d.
Большинство бактерий, способных к фотосинтезу, содержат бактериохлорофилл а , некоторые – бактериохлорофилл b, зелёные бактерии – c и d.
Хлорофилл достаточно эффективно поглощает лучистую энергию и передаёт её другим молекулам. Благодаря этому хлорофилл – единственное вещество на Земле, способное обеспечивать процесс фотосинтеза.
Пластидам, как и митохондриям, свойственна в определённой степени автономность внутри клетки. Они способны размножаться в основном путём деления.
Наряду с фотосинтезом в хлоропластах происходит синтез других веществ, таких как белки, липиды, некоторые витамины.
Благодаря наличию в пластидах ДНК, они играют определённую роль в передаче признаков по наследству (цитоплазматическая наследственность ).
Митохондрии – энергетические центры клетки
В цитоплазме большинства животных и растительных клеток содержатся достаточно большие овальные органеллы (0,2 – 7 мкм), покрытые двумя мембранами.
Митохондрии называют силовыми станциями клеток, потому что их основная функция – синтез АТФ. Митохондрии превращают энергию химических связей органических веществ на энергию фосфатных связей молекулы АТФ, которая является универсальным источником энергии осуществления для всех процессов жизнедеятельности клетки и целого организма. АТФ, синтезированная в митохондриях, свободно выходит в цитоплазму и дальше идёт к ядру и органеллам клетки, где используется её химическая энергия.
Митохондрии содержатся почти во всех эукариотических клетках, за исключением анаэробных простейших и эритроцитов. Они расположены в цитоплазме хаотично, но чаще их можно определить возле ядра или в местах с высокой потребностью в энергии.
Пример 2
В мышечных волокнах митохондрии расположены между миофибриллами.
Эти органеллы могут изменять свою структуру и форму, а также двигаться внутри клетки.
Количество органелл может изменяться от десятков до нескольких тысяч в зависимости от активности клетки.
Пример 3
В одной клетке печени млекопитающих содержится более 1000 митохондрий.
Структура митохондрий в некоторой мере отличается у различных типов клеток и тканей, но все митохондрии имеют принципиально одинаковое строение.
Образуются митохондрии путём деления. Во время деления клетки они более-менее равномерно распределяются между дочерними клетками.
Внешняя мембрана гладкая, не образует никаких складок и выростов, легко проницаема для многих органических молекул. Содержит ферменты, которые превращают вещества на реакционно способные субстраты. Участвует в образовании межмембранного пространства.
Внутренняя мембрана плохо проницаема для большинства веществ. Образует много выпячиваний внутрь матрикса – крист . Количество крист в митохондриях разных клеток неодинакова. Их может быть от нескольких десятков до нескольких сотен, причём особенно много их в митохондриях клеток, которые активно функционируют (мышечные). Содержит белки, которые участвуют в трёх важнейших процессах:
- ферменты, катализирующие окислительно-восстановительные реакции дыхательной цепи и транспорта электронов;
- специфические транспортные белки, участвующие в образовании катионов водорода в межмембранном пространстве;
- ферментативный комплекс АТФ-синтетазы, который синтезирует АТФ.
Матрикс – внутреннее пространство митохондрии, ограниченное внутренней мембраной. Он содержит сотни различных ферментов, которые участвуют в разрушении органических веществ вплоть до углекислого газа и воды. При этом освобождается энергия химических связей между атомами молекул, которая в дальнейшем превращается на энергию макроэргических связей в молекуле АТФ. В матриксе также есть рибосомы и молекулы митохондриальной ДНК.
Замечание 4
Благодаря ДНК и рибосомам самих митохондрий обеспечивается синтез белков, необходимых самой органелле, и которые в цитоплазме не образуются.
Митохондрии - это органеллы размером с бактерию (около 1 х 2 мкм). Митохондрии - неотъемлемая часть всех живых эукариотических клеток, обычно в клетке содержится около 2000 митохондрий, общий объем которых составляет до 25% от общего объема клетки. Форма, величина и их число постоянно меняются. Число митохондрий варьирует от нескольких десятков до сотен. Особенно их много в секреторных тканях растений.
Митохондрии независимо от их величины или формы имеют универсальное строение, их ультраструктура однообразна. Митохондрии ограничены двумя мембранами. Наружная митохондриальная мембрана отделяет ее от гиалоплазмы. Обычно она имеет ровные контуры, не образует впячиваний или складок. На нее приходится около 7% от площади всех клеточных мембран. Толщина этой мембраны около 7 нм, она не бывает связана ни с какими другими мембранами цитоплазмы и замкнута сама на себя, так что представляет собой мембранный мешок. Наружную мембрану от внутренней отделяет межмембранное пространство шириной около 10--20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс, или митоплазму. Характерной чертой внутренних мембран митохондрий является их способность образовывать многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист.
Рис. Схема общей организации митохондрии
1 -- внешняя мембрана; 2 -- внутренняя мембрана; 3 -- впячивания внутренней мембраны - кристы; 4 -- места впячиваний, вид с поверхности внутренней мембраны
Митохондрии способны к независимому от ядра синтезу своих белков на собственных рибосомах под контролем митохондриальной ДНК. Митохондрии образуются только путем деления.
Основная функция митохондрий состоит в обеспечении энергетических потребностей клетки путем дыхания. Богатые энергией молекулы АТФ синтезируются при реакции окислительного фосфорилирования. Энергия, запасаемая АТФ, получается в результате окисления в митохондриях различных энергетически богатых веществ, главным образом сахаров. Механизм окислительного фосфорилирования путем хемиосмотического сопряжения открыт в 1960 г. английским биохимиком П.Митчеллом
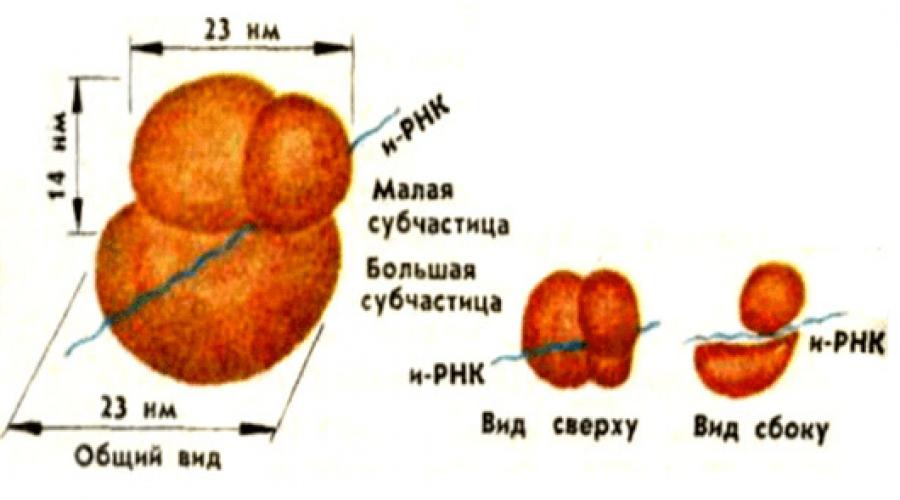
Основной функцией рибосом является трансляция, то есть синтез белков. На фотографиях, полученных с помощью электронного микроскопа, они выглядят округлыми тельцами диаметром 20 - 30 нм.
Каждая рибосома состоит из 2-х субъединиц неравных размеров, формы и строения. Субъединицы рибосом обозначают по величине коэффициентов седиментации (то есть осаждения при центрифугировании).

По-видимому, малая субъединица располагается поверх большой так, что между частицами сохраняется пространство («туннель»). Туннель используется для размещения м - РНК во время белкового синтеза.
Рибосомы представляют собой крупный рибонуклеопротеидный комплекс с молекулярной массой около 2,5 мДа, состоящий из рибосомных белков, молекул рРНК и ассоциированных с ними факторов трансляции. Рибосомы - немембранные органеллы, на которых происходит синтез белка в клетке. Они представляют собой сферические структуры с диаметром около 20 нм. Эти самые мелкие клеточные органеллы устроены чрезвычайно сложно. Ни одна молекула, входящая в состав рибосом, не повторяется дважды. Лучше других изучены рибосомы бактерии Е. coli.
История изучения строения рибосом насчитывает более полувека со времени их открытия, и краткое описание методов, использованных для этого, представляет отдельный интерес, поскольку эти методы используются или могут быть использованы для изучения не только рибосом, но и других сложных надмолекулярных комплексов.
Итак, к 1940 г. Альберт Клод (США) сумел выделить из эукариотических клеток цитоплазматические РНК-содержащие гранулы, гораздо меньшие, чем митохондрии и лизосомы (от 50 до 200 мкм в диаметре); позже он назвал их микросомами. Результаты химических анализов показали, что микросомы Клода были рибонуклеопротеидными комплексами. В дополнение к этому, цитохимические работы Т. Касперсона (Швеция) и Ж.Браше (Бельгия) продемонстрировали, что чем интенсивнее идет белковый синтез, тем больше обнаруживается РНК в цитоплазме.
В дальнейшем, некоторым исследователям удавалось выделять из клеток бактерий, животных и растений частицы, ещё более мелкие, чем микросомы. Электронная микроскопия и седиментационный анализ в ультрацентрифуге указывали, что частицы компактны, более или менее сферичны и гомогенны по размеру, имея диаметр 100-200 Ȧ (ангстрем) и обнаруживая резкие седиментационные границы с коэффициентами седиментации от 30-40S до 80-90S (S-коэффициент седиментации , или константа Сведберга, - отражает скорость осаждения каких-либо молекулярных комплексов при скоростном ультрацентрифугировании и зависит от молекулярного веса частиц и их плотности – компактности ). Пожалуй, первое ясное свидетельство, что такие частицы бактерий являются рибонуклеопротеидами было получено Г.К. Шахманом, А.Б. Парди и Р. Станиером (США) в 1952 г.
Улучшенная техника микротомии и электронной микроскопии ультратонких срезов животных клеток привела к выявлению однородных плотных гранул с диаметром около 150 Ȧ непосредственно в клетке. Электронно-микроскопические исследования Дж. Паладе (США) , проведенные в 1953-1955 гг., показали, что маленькие плотные гранулы в изобилии содержатся в цитоплазме животных клеток. Они видны либо присоединенными к мембране эндоплазматического ретикулума, либо свободно рассеяны в цитоплазме. Микросомы Клода оказались фрагментами эндоплазматического ретикулума с сидящими на них гранулами. Выяснилось, что эти «гранулы Паладе» являются рибонуклеопротеидными частицами и что они представляют основную массу цитоплазматической РНК, обеспечивающей белковый синтез.
Исследования функциональной роли рибосом шли параллельно с их обнаружением и структурным описанием. Первой убедительной демонстрацией того, что именно рибонуклеопротеидные частицы микросом ответственны за включение аминокислот в новосинтезированный белок, были эксперименты П. Замечника с сотрудниками (США), опубликованные в 1955 г. За этим последовали эксперименты из этой же лаборатории, показавшие, что свободные рибосомы не прикрепленные к мембранам эндоплазматического ретикулума, также включают аминокислоты и синтезируют белок, освобождающийся затем в растворимую фазу. Функции бактериальных рибосом были предметом интенсивных исследований группы Р.Б. Робертса (США); публикация К. МакКиллена, Р.Б. Робертса и Р.Дж. Бриттена в 1959 г. окончательно установила, что белки синтезируются в рибосомах и затем распределяются по другим частям бактериальной клетки.
Митохондрии являются органоидами всех эукариотических клеток. Они характеризуются обилием внутренних мембран. Две мембраны - внешняя и внутренняя - отделяют их от цитоплазмы. Мембраны образуют в митохондриях большие внутренние компартменты, в которых происходят реакции окислительного фосфорилирования. В результате этих процессов энергия реакций окисления преобразуется в энергию, заключенную в молекулах АТФ. При этом митохондрии исключительно эффективно используют для окисления сахара и жирные кислоты.
Митохондрии (греч. mitos-нить, chondros-зерно) занимают в эукариотических клетках значительную часть цитоплазмы. Подсчеты показывают, что на одну печеночную клетку приходится около тысячи митохондрий. Это примерно 20% общего объема цитоплазмы и около 30-35% общего количества белка в клетке. В ооцитах насчитывается до 300000 митохондрий, в гигантских амебах до 500000. В клетках зеленых растений митохондрий меньше, чем в клетках животных.
Митохондрии были описаны еще в конце прошлого века, так как их размеры довольно велики, то они сопоставимы с размерами бактериальной клетки, и хорошо различимы с помощью светового микроскопа. В типичном случае митохондрии представляют собой цилиндр диаметром 0,5 мкм и длиной до 1 мкм. Однако у разных организмов длина митохондрий колеблется в значительных пределах от 7 до 10 мкм. В клетках дрожжей, клетках мышечной ткани, у трипаносом присутствуют разветвленные паукообразные митохондрии. Они обладают достаточно высокой плотностью, благодаря чему их можно наблюдать в живых клетках. Такие наблюдения с помощью микрокиносьемки показывают, что форма митохондрий в живых клетках весьма изменчива, это необыкновенно подвижные и пластичные органоиды. В течение минуты они могут изменить свою цилиндрическую форму 15-20 раз, принимая вид пузырьков, гантелей, теннисной ракетки, они могут изгибаться и выпрямляться.
Локализация митохондрий в клетках определяется двумя факторами. Во-первых, она зависит от расположения других органоидов и включений. В растительных дифференцированных клетках митохондрии отодвигаются к периферии клетки центральной вакуолью, в клетках меристемы они располагаются более-менее равномерно. В делящихся клетках митохондрии располагаются также периферически, их вытесняет веретено деления. Ориентацию митохондрий могут определять микротрубочки цитоплазмы. Во-вторых, митохондрии скапливаются в энергозависимых участках клетки. В скелетных мышцах - между миофибрилл, в сперматозоидах плотно обвивают жгутик, у простейших, снабженных ресничками, митохондрии лежат у основания ресночек под плазматической мембраной. В нервных клетках - около синапсов, где происходит передача нервных импульсов. В секреторных клетках митохондрии связаны с зонами шероховатой ЭПС.
Реальная возможность понять тонкое строение митохондрий и их функций появилась только после 1948 г., когда были разработаны методы выделения митохондрий из клеток и началось их биохимическое исследование. Каждая митохондрия окружена двумя высокоспециализированными мембранами, играющими основную роль в ее работе. Эти мембраны образуют два изолированных митохондриальных компартмента - межмембранное пространство и внутренний матрикс. Внутренняя мембрана образует многочисленные кристы, увеличивающие ее общую поверхность.
Матрикс содержит высококонцентрированную смесь сотен различных ферментов, необходимых для окисления пирувата, жирных кислот, и ферментов цикла лимонной кислоты. 67% всего белка митохондрий приходится на матрикс. В матриксе содержится собственная ДНК, представленная несколькими идентичными молекулами и близка к бактериальной по составу нуклеотидов, кроме того она тоже кольцевая как у бактерий. Матрикс митохондрий включает и специфические митохондриальные рибосомы. По своим свойствам они также близки к бактериальным (70S).
Присутствие ДНК, рибосом и ферментов, участвующих в работе митохондриального генома, говорит о некоторой автономности митохондрий.
В митохондриях происходит синтез АТФ на основе окисления органических субстратов и фосфорилирования АДФ. Высвобождение энергии при аэробном окислении пищи называется дыханием.
Две международные команды ученых исследовали структуру митохондриальных рибосом с помощью криоэлектронной микроскопии. Этот метод позволяет увидеть структурные элементы с высочайшим разрешением. Новые сведения дали возможность сравнить детали строения цитоплазматических и митохондриальных рибосом. Как выяснилось, митохондриальные рибосомы высокоспециализированы и сильно отличаются и от цитоплазматических аналогов, и от бактериальных рибосом.
Хорошо известно, что митохондрии - это бывшие альфа-протеобактерии , которые примерно полтора миллиарда лет назад стали симбионтами клеток архей или каких-то иных клеток. Там они взяли на себя функцию энергетических снабженцев, усовершенствовав биохимический конвейер по производству АТФ - главной энергетической молекулы клетки. Зато другие функции жизнеобеспечения за них стала выполнять хозяйская клетка со своим ядром и регуляторами. Об оставленной свободной жизни в митохондриях напоминает присутствие мембран, собственной ДНК и рибосом, необходимых для изготовления небольшого набора митохондриальных белков. Все эти элементы высоко специализированы, так как нацелены, в отличие от всех остальных частей клетки, на выполнение только двух функций - производства АТФ и собственного размножения в стабильных внутриклеточных условиях. Поэтому изучение любого из этих элементов дает представление о процессах эволюционной специализации. Это касается в том числе и рибосом , хотя, казалось бы, эта клеточная машинка для синтеза белков универсальна, в ее работе уже ничего не убавить и не прибавить. Но оказалось, что это не так: митохондриальные рибосомы отличаются и от клеточных соседей, и от предковых рибосом альфа-протеобактерий. Это выяснили специалисты из в Цюрихе и Цюрихского университета . Также интересную работу на эту тему выполнили ученые из лаборатории молекулярной биологии Совета медицинских исследований в Кембридже.
Эти группы использовали криоэлектронную микроскопию (Cryo-electron microscopy), позволяющую реконструировать трехмерное изображение объектов с разрешением 3,4–3,8 ангстрем. При подготовке препаратов для криоэлектронной микроскопии не используются вспомогательные материалы для срезов, изменяющие структуру мелких клеточных включений. До сих пор, однако, разрешающая способность криоэлектронной микроскопии была не очень высока, и только теперь она усовершенствовалась до уровня высокоточной рентгеновской кристаллографии (которая позволяет устанавливать атомарную структуру вещества, см.: X-ray crystallography). С помощью этой техники оказалось возможным рассмотреть в подробностях различные субъединицы миторибосом (митохондриальных рибосом), соотнести биохимические и структурные различия с таковыми цитоплазматических рибосом.
Рибосомы - это комплексы белков и РНК, белки в рибосомах в основном являются рибозимами , что указывает на их подчиненную каталитическую роль в этом тандеме. В миторибосомах у млекопитающих (были изучены человеческие и свиные клетки) содержится меньше РНК и, соответственно, больше белков. В некоторых случаях белки заменяют потерянные части РНК, они покрывают практически всю рибосому, вероятно чтобы стабилизировать неустойчивую структуру РНК и защитить комплексы от окисления. Около половины миторибосомных белков специфичны: таких нет ни в цитоплазматических рибосомах, ни у родственных бактериальных рибосом. Так, у человека имеется 80 миторибосомных белков, из которых 36 специфичны. Одно из любопытных структурных различий, как выяснилось, такое: важный функциональный элемент рибосомы - малая субъединица 5S рРНК (5S ribosomal RNA) - замещена в митохондриях на тРНК валина. Эта замена особенно важна в свете дискуссий о природе 5S рРНК (см.: Г. М. Гонгадзе, 2011. 5S рРНК и рибосома), ее подозрительном сходстве с тРНК и возможном происхождении одной молекулы от другой (причем пока не ясно, какая именно от какой произошла).
Как сказались эти трансформации на работе миторибосом? Ученые предполагают, что именно они позволили миторибосомам стать специалистами по производству гидрофобных белков; и даже более - локализовать это производство на мембранах митохондрий. Нашлись специальные комплексы, которые прикрепляют рибосомы к мембранам митохондрий; нашлись особые белки, которые обеспечивают специфическую элонгацию; нашлись белки, которые занимаются распознаванием и прикреплением мРНК к миторибосоме. Все они различаются с функциональными аналогами цитоплазаматических рибосом. В особенности это касается инициации связывания мРНК с рибосомой - последней из перечисленных функций. То место, куда между двумя субъединицами входит нить матричной РНК, устроено у миторибосомы совершенно не так, как в цитоплазматической рибосоме. Именно из-за ее специфики ученые не могли наладить синтез митохондриальных белков in vitro, хотя цитоплазматические рибосомы работают в искусственных условиях уже более полувека. Теперь можно начать экспериментировать и с митохондриальными рибосомами.
Особенности миторибосомальных белков обуславливают иное устройство взаимодействия между малой и большой субъединицами. Из-за этого меняются конформационные движения и повороты этих субъединиц при связывании с тРНК и продвижении мРНК и синтезирующейся аминокислотной цепочки. Иными словами, механика работы миторибосомы при синтезе белковой нити отличается от канонической цитоплазматической рибосомы.
Обе команды исследователей подчеркивают, что обнаруженная специфика миторибосом объясняет побочные действия нескольких классов лекарств. Это означает, что структуру новых лекарств нужно немного изменить, чтобы устранить вредные последствия. Теперь стало понятно, куда смотреть и что менять. Хотя бы поэтому данная работа с миторибосомами актуальна. Хотя теоретический интерес специфики миторибосом гораздо шире: ведь известно, что миторибосомы сильно различаются у разных видов, гораздо сильнее, чем цитоплазматические рибосомы. Траектории изменений у разных видов покажут особенности энергетического обмена и пути его приспособления к разным модификациям.
Источники:
1) A. Amunts, A. Brown, J. Toots, S. H. W. Scheres, V. Ramakrishnan. The structure of the human mitochondrial ribosome // Science
. 2015. V. 348. P. 95–98.
2) A. Amunts, A. Brown, X. Bai, J. L. Llácer, T. Hussain, P. Emsley, F. Long, G. Murshudov, S. H. W. Scheres, V. Ramakrishnan. Structure of the Yeast Mitochondrial Large Ribosomal Subunit // Science
. 2014. V. 343. P. 1485–1489.
3) B. J. Greber, P. Bieri, M. Leibundgut, A. Leitner, R. Aebersold, D. Boehringer, N. Ban. The complete structure of the 55S mammalian mitochondrial ribosome // Science
. 2015. V. 348. P. 303–307.
4) R. Beckmann, J. M. Herrmann. Mitoribosome Oddities // Science
. 2015. V. 348. P. 288–289.
Елена Наймарк